Менструальный цикл
Второй уровень — яичники, в которых непрерывно, начиная с внутриутробного периода и заканчивая глубокой старостью женщины, происходят рост и созревание фолликулов (фолликулогенез), а также взаимосвязанный с этим процессом синтез стероидов (стероидогенез). Фолликулогенез (рис. 6--20) подразумевает собой цикл развития фолликула от примордиального до предовуляторного, с последующей овуляцией и образованием жёлтого тела.
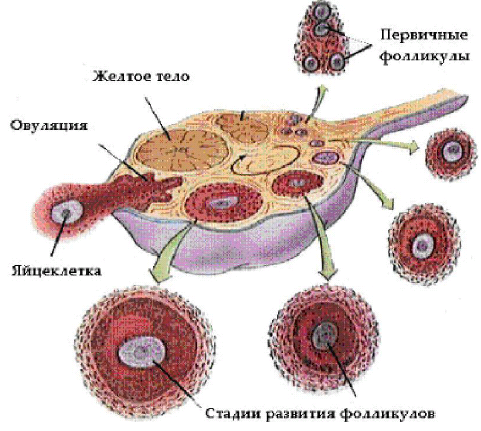
Рис. 6-20. Схема развития фолликула.
У взрослой женщины ежемесячно созревают примерно 20 фолликулов, что биологически необходимо для быстрой регенерации эндометрия под воздействием суммарного количества синтезируемых ими эстрогенов. Однако, поскольку для человека типична беременность одним плодом, на протяжении менструального цикла овулирует только один фолликул, который получает наиболее адекватное кровоснабжение и имеет оптимальную чувствительность к рецепторам гонадотропинов. Это позволяет ему получить самую высокую митотическую активность, накопить максимальное количество фолликулостимулирующего гормона (ФСГ) в фолликулярной жидкости и осуществить самый интенсивный синтез эстрадиола и ингибина по сравнению со всеми другими растущими фолликулами. В результате они претерпевают атретические изменения на различных этапах созревания, а лидирующий фолликул
становится доминантным и проходит полный цикл развития. Его размер к моменту овуляции в результате увеличения количества клеток гранулёзы с 0,5´106 до 50´106 возрастает с 2 мм (примордиальный фолликул) до 20–22 мм, объём фолликулярной жидкости соответственно увеличивается в 100 раз.
Примордиальный фолликул состоит из яйцеклетки, окружённой одним рядом уплощённых клеток эпителия, выстилающих фолликул изнутри. В процессе созревания фолликула яйцеклетка увеличивается, клетки фолликулярного эпителия размножаются и округляются, образуется зернистый слой фолликула (strarum granulosum). В гранулёзных клетках зреющего фолликула есть рецепторы к гонадотропинам, регулирующим процессы фоллиуо и стероидогенеза. В толще зернистой оболочки в результате секреции и распада клеток фолликулярного эпителия и транссудата из кровеносных сосудов появляется жидкость. Яйцеклетка оттесняется жидкостью к периферии, окружается 17–50 рядами клеток гранулёзы. Возникает яйценосный холмик (cumulus oophorus). Строма вокруг зреющего фолликула дифференцируется на наружное (tunica externa thecae folliculi) и внутреннее покрытие фолликула (tunica interna thecae folliculi). Когда фолликулярная жидкость начинает полностью окружать ооцит, зреющий фолликул превращается в зрелый (граафов пузырёк). В фолликулярной жидкости резко увеличивается содержание эстрадиола и ФСГ. Пропорциональный подъём концентрации E2 в периферической крови включает механизм обратной связи и стимулирует выброс лютеинизирующего гормона (ЛГ) и разрыв стенки фолликула ЛГ (овуляцию) в предварительно подготовленной области (стигме). Изменения в стенке фолликула (истончение и потенциальная возможность разрыва) обеспечиваются повышением активности фермента коллагеназы и протеолитических ферментов фолликулярной жидкости, а также ПГ-F2a и ПГ-E2, окситоцина и релаксина. На месте разорвавшегося фолликула образуется жёлтое тело, клетки которого секретируют прогестерон, эстрадиол и андрогены. Полноценное жёлтое тело образуется только тогда, когда в предовуляторном фолликуле содержится достаточное количество гранулёзных клеток с высоким содержанием рецепторов к ЛГ. Стероидные гормоны продуцируются клетками гранулёзы, клетками thecae folliculi interna и, в меньшей степени, theca folliculi externa. Клетки гранулёзы и тека-клетки преимущественно участвуют в синтезе эстрогенов и прогестерона, а клетки theca folliculi externa — в синтезе андрогенов. Исходный материал для всех стероидных гормонов — холестерол, образующийся из ацетата или липопротеидов низкой плотности, который поступает в яичник с током крови. Андрогены синтезируются в тека-клетках под влиянием ЛГ и с током крови попадают в гранулёзные клетки. Конечные этапы синтеза (превращение андрогенов в эстрогены) происходят под влиянием ферментов ароматаз, образующихся при участии ФСГ. В клетках гранулёзы образуется белковый гормон — ингибин, тормозящий выделение ФСГ. В фолликулярной жидкости, жёлтом теле, матке и маточных трубах обнаружен окситоцин, который секретируется яичником и оказывает лютеолитическое действие, способствуя регрессу жёлтого тела. Вне беременности в клетках гранулёзы и жёлтого тела в очень небольших количествах вырабатывается релаксин, в жёлтом теле беременности его содержание возрастает во много раз. Релаксин оказывает токолитическое действие на матку и способствует овуляции. Третий уровень — передняя доля гипофиза (аденогипофиз). Здесь секретируются гормоны, тропные для эндокринных желёз:
· гонадотропины (ФСГ, ЛГ, пролактин);
· тиреотропный гормон (ТТГ);
· соматотропный гормон;
· адренокортикотропный гормон (АКТГ);
· меланоцитстимулирующий гормон.
ФСГ и ЛГ — гликопротеиды, пролактин — полипептид.
Железамишень для ФСГ и ЛГ — яичник, синхронный выброс ЛГ и ФСГ способ-ствует овуляции. ФСГ стимулирует рост фолликула, пролиферацию клеток гранулёзы, образование рецепторов ЛГ на поверхности клеток гранулёзы. ЛГ увеличивает образование андрогенов в тека-клетках и синтез прогестерона в лютеинизированных клетках гранулёзы после овуляции. Основная роль ПРЛ — стимуляция роста молочных желёз и регуляция лактации. Он оказывает гипотензивное действие, даёт жиромобилизующий эффект, контролирует деятельность жёлтого тела. Повышение уровня ПРЛ тормозит фолликулогенез и стероидогенез в яичниках. Четвёртый уровень репродуктивной системы — гипофизотропная зона гипоталамуса (рис. 6--21), её вентромедиальные, дорсомедиальные и аркуатные ядра, обладающие нейросекреторной активностью — способностью синтезировать либерины и статины (рилизинг-гормоны).
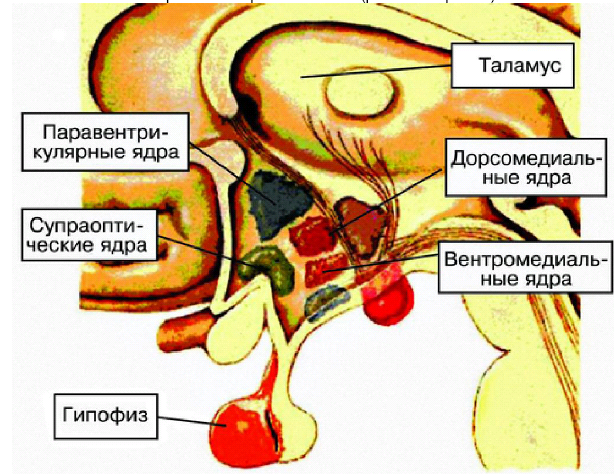
Рис. 6-21. Гипофизотропная зона гипоталамуса.
Выделен, синтезирован и описан ГнРГ (люлиберин), который стимулирует выделение клетками передней доли гипофиза ЛГ и, частично, ФСГ. ГнРГ из аркуатных ядер по аксонам нервных клеток попадает в терминальные окончания, тесно соприкасающиеся с капиллярами медиальной возвышенности гипоталамуса, которые формируют портальную кровеносную систему, объединяющую гипофиз и гипоталамус. Её особенность — возможность тока крови в обе стороны, что важно для осуществления механизма обратной связи. Нейросекрет гипоталамуса оказывает биологическое воздействие на организм различными путями. Основной путь — парагипофизарный — через вены, впадающие в синусы твёрдой мозговой оболочки, а оттуда в ток крови.
Трансгипофизарный путь — через систему воротной вены к передней доле гипофиза. Обратное влияние на гипоталамус половых стероидов осуществляется через вертебральные артерии. Секреция ГнРГ генетически запрограммирована и происходит в определённом пульсирующем ритме с частотой примерно один раз в час. Этот ритм получил название цирхорального (часового). Он формируется в пубертатном возрасте и считается показателем зрелости нейросекреторных структур гипоталамуса. Цирхоральная секреция ГнРГ запускает гипоталамо-гипофизарно- яичниковую систему. Под влиянием ГнРГ происходит выделение ЛГ и ФСГ из клеток передней доли гипофиза. В модуляции пульсации ГнРГ играет роль эстрадиол. Величина выбросов ГнРГ в предовуляторный период (на фоне максимального выделения эстрадиола) значительно выше, чем в раннюю фолликулиновую и лютеиновую фазы. Частота выбросов остаётся прежней. В дофаминергических нейронах аркуатного ядра гипоталамуса есть рецепторы к эстрадиолу. Основная роль в регуляции выделения пролактина принадлежит дофаминергическим структурам гипоталамуса. Дофамин тормозит выделение пролактина из гипофиза. Пятый уровень регуляции менструального цикла — надгипоталамические церебральные структуры. Воспринимая тимпульсы из внешней среды и от интерорецепторов, они передают их через систему передатчиков нервных импульсов (нейротрансмиттеров) в нейросекреторные ядра гипоталамуса. В эксперименте показано, что в регуляции функции гипоталамических нейронов, секретирующих ГнРГ, ведущая роль принадлежит дофамину, норадреналину и серотонину. Функцию нейротрансмиттеров выполняют нейропептиды морфиноподобного действия (опиоидные пептиды) — эндорфины и энкефалины, регулирующие функцию гипофиза. Эндорфины подавляют секрецию ЛГ, а их антагонист — налоксон — приводит к резкому повышению секреции ГнРГ. Считают, что эффект опиоидов осуществляется в результате изменения содержания дофамина (эндорфины снижают его синтез, вследствие чего стимулируются секреция и выделение пролактина). В регуляции менструального цикла участвует кора большого мозга. Существуют данные об участии амигдалоидных ядер и лимбической системы в нейрогуморальной регуляции менструального цикла. Электрическое раздражение амигдалоидного ядра (в толще больших полушарий) вызывает в эксперименте овуляцию. Нарушения овуляции наблюдают при стрессовых ситуациях, перемене климата, ритма работы. Нарушения менструального цикла реализуются через изменение синтеза и потребления нейротрансмиттеров в нейронах мозга. Таким образом, репродуктивная система представляет собой суперсистему, функциональное состояние которой определяется обратной связью входящих в неё подсистем, как отрицательной, так и положительной (овуляция).
